



Recent announcements herald impressive computational milestones: the JUPITER supercomputer aims to simulate 20 billion neurons with 100 trillion connections, matching the human cerebral cortex in scale. The technical achievement is impressive and undeniable. Yet it prompts fundamental questions about the relationship between computational scale and scientific understanding. As both artificial intelligence and neuroscience grapple with the limits of “scaling is all you need”, we have an opportunity to chart a more productive path, one that prioritizes mechanistic insight over numerical magnitude.
Lessons from AI’s Scaling Plateau
The AI community recently confronted sobering realities about scaling. As Gary Marcus documented in “Scale Is All You Need is Dead,” simply increasing model size, data, and compute proved insufficient for artificial general intelligence (AGI). At NeurIPS 2025, leading researchers acknowledged persistent challenges with reliability, reasoning, hallucination, and generalization—despite unprecedented investments. Multiple studies from MIT, McKinsey, and BCG found that 95% of companies report disappointing returns on generative AI investments.
This doesn’t invalidate the progress. Large language models have achieved remarkable capabilities and their usefulness and impact is undeniable. But the dominant argument in the past few years, that AGI is just a matter of scaling the existing architectures, is now provably wrong: scale amplifies what’s present but doesn’t create what’s absent. As François Chollet and Yann LeCun argue, we need architectural innovations that capture fundamental computational principles, not just bigger versions of existing approaches.
The Allure (and limitations) of Brain-Scale Simulation
Computational neuroscience faces analogous questions, but with an added layer of complexity. Current brain-scale simulation efforts primarily use simplified point neuron models—typically leaky integrate-and-fire (LIF) neurons connected via simplified, static synapses. Recent coverage of these efforts presents them as breakthroughs that will “offer unprecedented insights into how our brains work.”
The reality is more nuanced.
The argument for scale echoes recent AI optimism: “Large language models have shown that larger systems contain features simply not present in smaller ones,” suggests Markus Diesmann. “We know now that large networks can do qualitatively different things than small ones.”
This observation is correct—but also insufficient. Let me explain why through four interconnected arguments.
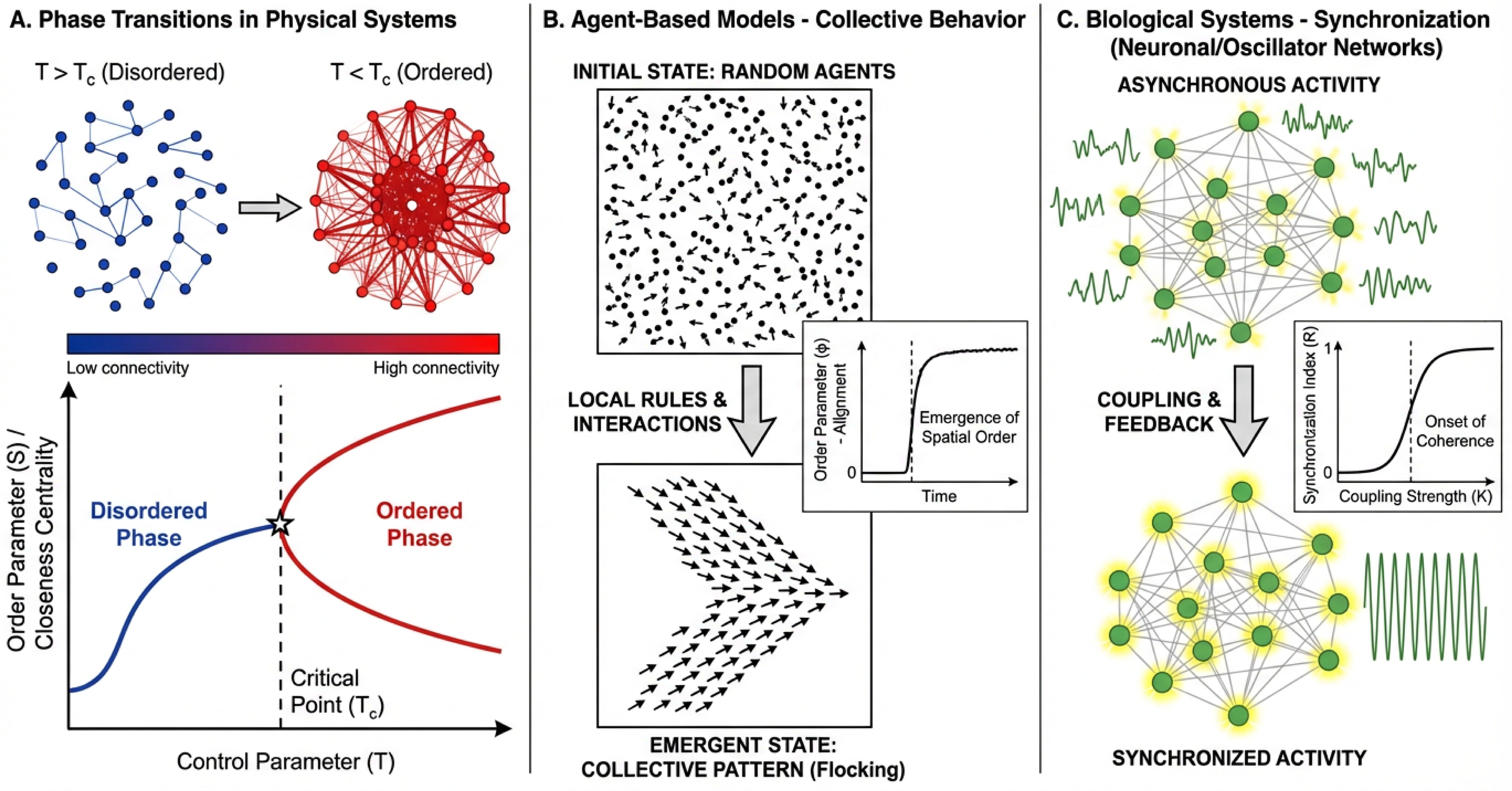
I. Emergence is Expected, Not Explanatory
First, the appearance of qualitatively different behaviors at scale is neither surprising nor, by itself, scientifically revealing. Nonlinear dynamical systems naturally display emergent properties—the whole becomes genuinely more than the sum of its parts. This is a fundamental characteristic of complex systems, well-established in physics, chemistry, and mathematics.
Recent analyses of emergent abilities in large language models note that “both chaotic systems and deep neural networks are characterized by high dimensionality and inherent non-linearity, which are prerequisites for complex and sensitive behavior and emergence.” The observation that scale produces emergence is an empirical fact—but one with limited explanatory power unless we understand the mechanisms generating those emergent phenomena. I had a professor that said he didn’t like the term “emergence” because he equated it to “we have no idea what is causing this”.
In brain-scale simulations, observing network-level dynamics different from the sum of single-neuron behavior tells us something happened at scale. It doesn’t tell us why. It doesn’t tell us if those dynamics match biological reality. It doesn’t tell us which features are computationally essential versus artifacts of the particular implementation choices.
The real question is whether our models capture the mechanisms that generate the emergent phenomena we care about.
II. What Simplification Strips Away: The Descriptive Adequacy Problem
This brings us to the core problem: descriptive adequacy—whether a model simplifies away the computational elements that generate the phenomena we aim to explain.
As I have previously argued, there’s a fundamental tension in modeling: “If a model simplifies away the core computational elements of the system, our ability to account for its operations is lost.” The challenge isn’t avoiding simplification—all models simplify—but rather asking “what is the cost of simplification?”
Current brain-scale models use point neurons (LIF models) and point-current synapses that reduce each neuron to a summation device firing when inputs exceed threshold. These models omit crucial biological mechanisms that aren’t implementation details but computational substrates. Stripping them away removes a fundamental computational layer which may be causally responsible for the observed dynamics and computational capabilities (and, consequently, cognitive and behavioral phenotypes), i.e., the emergent phenomena we care about. For instance:
Dendritic computation: Real neurons aren’t point integrators. London and Häusser’s seminal work demonstrated that dendrites perform active computation—individual branches execute nonlinear integration, detect coincident inputs, and generate local spikes, creating compartmentalized subnetworks within single neurons. Recent work confirms these mechanisms are essential for in vivo neural computations and behavior and we can now efficiently model them.
Ion channel and Receptor diversity: Neurons express hundreds of ion channel types with distinct kinetics, voltage dependencies, ligand dependencies and spatial distributions, optimized for energy efficiency and computational flexibility. These channels enable neurons to implement multiple computational modes that threshold models cannot capture. Additionally, the properties and distribution of these molecular components is adaptive and subjected to activity-dependent modulation.
Neuromodulation: Computational studies of neuromodulation reveal how dopamine, serotonin, and acetylcholine dynamically reconfigure network properties, enabling the same circuit to implement different computations based on behavioral context. Networks of point neurons and static current synapses lack these state-dependent dynamics entirely and extending them in that direction is far from trivial as they lack the relevant processes and variables in the first place.
Synaptic plasticity mechanisms: Multi-scale studies show learning occurs through complex biochemical cascades operating across timescales ranging from milliseconds to hours—crucial for understanding memory, development, and adaptation. Suffice it to say that until we understand the molecular and cellular, mechanistic bases, of plasticity and appropriately incorporate them (at the relevant level of abstraction), we will not capture learning, memory and a host of cognitive processes.
Brain-scale simulations with oversimplified components become anatomically constrained activity propagation models: simulations that respect connectivity structure but treat neurons as simple relays, lacking the biophysical features that generate actual computation. At this level of abstraction, the model’s relationship to the brain is primarily architectural, not functional.
III. What Brain-Scale Models Can Tell Us
This isn’t to say structurally constrained models are worthless. But we should be clear about what they can—and cannot—reveal.
In 2014, Potjans and Diesmann published their landmark cortical microcircuit model: approximately 80,000 neurons organized into cortical layers 2/3, 4, 5, and 6 (E/I sub-populations in each layer), connected according to empirically derived connectivity statistics. The model used simple LIF neurons and demonstrated that “the observed cell-type and layer specificity of in-vivo firing rates is largely explained by the specificity in the number of connections between cortical subpopulations.”
This was a genuine insight—but notice what it revealed: how connectivity architecture constrains activity flow and firing rate distributions. The model succeeded precisely because its scientific question matched its level of description. It didn’t attempt to explain computational processes, learning mechanisms, or cognitive function. It asked: given this connectivity structure, what activity patterns emerge?
Current brain-scale simulations occupy similar conceptual territory. The primary insights concern signal propagation and information flow through anatomically defined networks. This is valuable for specific (and perfectly legitimate) questions, but one has to keep in mind that the observed dynamics will be a consequence of the chosen formalisms to model the network’s nodes and edges. And, if these building blocks differ fundamentally from the corresponding biological system, the range of scientific questions we can explore is narrow. In fact, if we try to quantify information processing capacity of prominent cortical circuit models designed with similar priorities in mind, we realize their limitations as models of the biological system.
Brain-scale models will likely reveal insights about large-scale signal propagation, network topology effects, and spatiotemporal dynamics across interconnected brain regions. Understanding these phenomena matters. But these questions are fundamentally different from understanding how neural circuits compute, how learning occurs, how cognition emerges or how mechanistic dysfunctions yield clinical phenotypes.
For the latter questions—the ones that motivated building brain-scale models in the first place—simplification may have crossed into inadequacy.
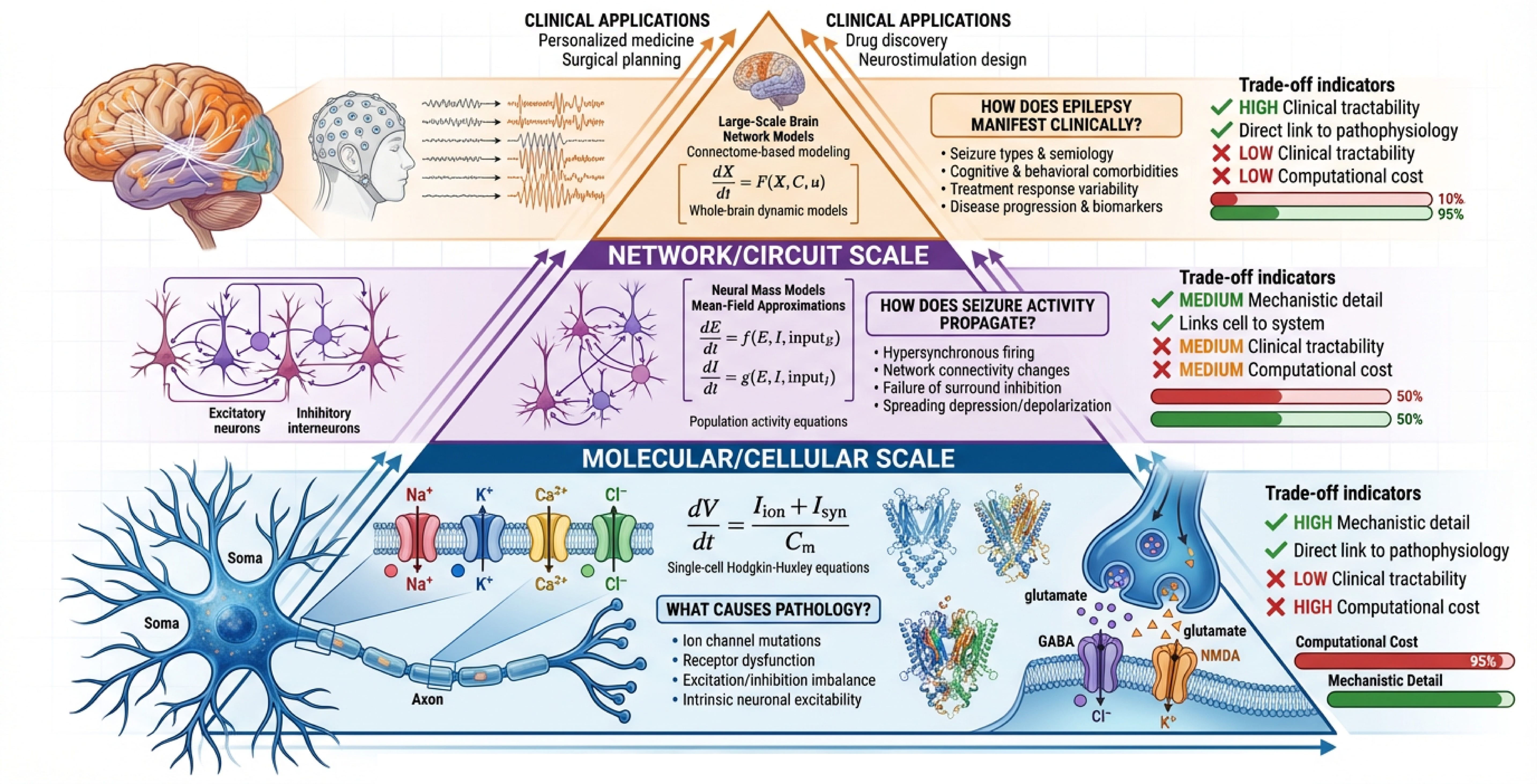
IV. Disease Modeling: When Simpler is More Insightful
This brings us to a crucial paradox: for the very applications where brain-scale models are promoted most heavily—disease modeling—the supposed advantage of scale may be illusory.
The translational promise of brain-scale simulation deserves special attention. Neurological and psychiatric disorders—epilepsy, Parkinson’s disease, Alzheimer’s disease, schizophrenia—arise from specific molecular and cellular pathologies. For disease modeling, biological detail isn’t optional; it’s essential.
Yet here’s the paradox: for many disease applications, simpler population-level models may be more insightful than detailed brain-scale simulations.
Consider epilepsy, one of the flagship applications for brain-scale modeling. Epileptic seizures manifest as mesoscopic and macroscopic phenomena—abnormal synchronization across neuronal populations visible in EEG and MEG. The pathology often involves altered ion channel kinetics, synaptic receptor dysfunction, or circuit-level excitability changes.
Neural mass and neural field models—coarse-grained population descriptors—have proven remarkably successful for epilepsy research. Recent work shows that “mean-field models are often preferred over the more detailed models since they have fewer parameters and, thus, simplify the study of transitions from interictal to ictal states.” These models successfully capture seizure initiation, propagation, and termination while remaining computationally tractable for personalization and intervention testing.
Crucially, patient-specific “digital twin” approaches for epilepsy surgery planning use neural mass models, not detailed spiking networks. They work: retrospective validation shows these models can localize epileptogenic zones and predict surgical outcomes. The reason they work is that they operate at the right scale for the phenomenon—population-level dynamics where seizures actually manifest.
This doesn’t mean microscopic detail is irrelevant. Ion channel mutations cause epilepsy; understanding those mechanisms requires biophysically detailed models. But the optimal modeling strategy is multiscale, hybrid and mechanism-specific—detailed biophysical models for understanding molecular pathology, population models for understanding network-level seizure dynamics, and principled links between scales.
As emphasized in recent reviews, “although biophysically explicit modeling is the primary technique to look into the role played by experimentally inaccessible variables in epilepsy, the usefulness of detailed biophysical models is limited by constraints in computational power, uncertainties in detailed knowledge of neuronal systems, and the required simplification for numerical analysis.”
Simply scaling up LIF networks to brain-scale doesn’t solve this, it doesn’t even begin to address the problem. You get neither the mechanistic precision of targeted biophysical models nor the analytical tractability of population models. For many questions, you occupy an awkward middle ground: too detailed for efficient exploration, too simplified for mechanistic insight.
A Constructive Path Forward
This critique is not aimed at dismissing large-scale simulation but rather redirecting computational resources (and funding) toward maximum scientific insight. Several productive directions emerge:
Multiscale Integration with Purpose: Rather than uniform brain-scale detail, build hybrid models that deploy detail where mechanistically necessary. Recent reviews demonstrate the power of integrating molecular mechanisms, detailed microcircuit models, and large-scale dynamics—but only where each level informs specific questions. The key insight: extract “a broad range of behavior” by connecting different organizational levels, not by simulating everything everywhere all at once.
Mechanism-Specific Depth: Instead of simulating the entire brain with uniform simplification, target specific phenomena with appropriate biological detail. For working memory, include mechanisms relevant for persistent activity. For sensory processing, capture dendritic integration of feedforward and feedback inputs. As Schirner et al. showed, brain network models “support multi-scale neurophysiological inference” by carefully matching model complexity to scientific questions.
Validation Frameworks: The Human Brain Project’s recent review emphasized that modeling efforts need rigorous validation frameworks. Models should make testable predictions at multiple scales—single-neuron responses to network dynamics to behavioral outputs—and be systematically compared against experimental data. Without validation, scale is just spectacle.
Hybrid Computational Approaches: Recent innovations combine biophysical simulations with machine learning to bridge scales efficiently. These hybrid methods use detailed models to capture mechanisms while employing statistical approaches to scale up, balancing biological realism with computational tractability.
Component-Based Modeling: Move away from monolithic simulations toward modular frameworks where validated components—channel models, plasticity rules, circuit motifs—can be composed and tested systematically. This increases model reusability and facilitates systematic comparison.
Bridging Neuroscience and AI: Principled Abstraction
Both fields may benefit from reconvergence—but not through naive brain-simulation-as-AI or AI-as-brain analogies. As recent reviews on bio-inspired AI argue, incorporating biological principles (not necessarily implementation details) could guide more robust artificial systems.
The key is principled abstraction: identifying which biological features are computationally essential for the phenomena under investigation and which can be safely simplified. This requires theory to guide both models and experiments, not just assembling larger networks and hoping insights emerge.
The Real Value Proposition
Let’s be clear about what brain-scale simulation can and cannot deliver:
What it can deliver:
Insights into signal propagation through anatomically constrained networks
Understanding how structural connectivity shapes dynamics and oscillations
Testbeds for interventions (lesions, stimulation) at network scale
Benchmarks for computational methods and infrastructure
Integration of multiple data modalities (anatomy, connectivity, physiology)
What it struggles to deliver:
Understanding of how neural circuits compute (requires biophysical mechanisms)
Mechanistic accounts of learning and plasticity (requires molecular-, cellular- and circuit-level details)
Predictive models of cognition and behavior (requires computational principles)
Disease mechanisms rooted in molecular/cellular pathology (too abstracted)
Cost-effective insight compared to recording from actual brains (at this level of simplification, empirical neuroscience is often more informative)
The uncomfortable truth: at brain scale with point neurons, you’re studying a system whose primary relationship to the brain is anatomical connectivity. And connectivity alone doesn’t determine what computations are possible, nor does it provide mechanistic or principled causal explanations. It cannot compute in the ways real brains compute (or at all, in most instances) because it lacks the biophysical substrates for computation. It cannot learn in the ways real brains learn (or at all, in most instances) because it lacks the relevant plasticity mechanisms. It cannot inform disease mechanisms because it lacks the molecular and cellular features where pathology occurs.
Conclusion: Scale with Insight
Brain-scale simulation and exascale computing represent remarkable engineering feats. The question isn’t whether we can simulate 20 billion neurons—we can—but whether doing so advances understanding of neural computation, cognition, and disease.
The field’s history offers guidance. Hodgkin and Huxley’s model succeeded not through comprehensive detail but through principled reduction: carefully isolating mechanisms and deriving equations grounded in experimental measurements. Their 26-parameter model, with almost no unconstrained variables, explained action potential generation and predicted novel phenomena.
Today’s computational power enables ambitious synthesis—but power without principle risks generating heat instead of light. As Gerstner et al. emphasized, models should be “aids for understanding,” not facsimiles of reality.
The opportunity before us: multiscale integration connecting molecules to circuits to behavior; mechanism-specific models targeting phenomena with appropriate detail; validation frameworks ensuring predictions meet empirical reality; hybrid approaches balancing realism and tractability; modular components enabling systematic exploration.
This requires not abandoning large-scale simulation but directing computational resources and funding toward biologically meaningful questions where scale serves scientific discovery—not as an end in itself, but as a means to mechanistic understanding.
Scaling is not all we need, not in AI, not in neuroscience. When combined with biological insight, mechanistic understanding, and rigorous validation, computational approaches can genuinely illuminate how neural systems generate cognition and behavior.
But scale without mechanism is just spectacle.
The future of computational neuroscience lies not in simulating more neurons, but in understanding which neurons, which mechanisms, and which scales matter for the questions we’re asking. That’s where genuine insight emerges.
Key References:
Diesmann, M. et al. (2024). Brain-scale spiking neural networks on exascale systems. arXiv.
Potjans, T.C. & Diesmann, M. (2014). The cell-type specific cortical microcircuit. Cerebral Cortex, 24(3), 785–806.
Ranjan, R. & Prescott, S. (2016). Is realistic neuronal modeling realistic? J Physiology, 594(22).
London, M. & Häusser, M. (2005). Dendritic computation. Ann Rev Neurosci, 28.
Cakan, C. & Obermayer, K. (2020). Biophysically grounded mean-field models. PLOS Comp Biol, 16(7).
Dura-Bernal, S. et al. (2024). Large-scale mechanistic models with biophysically detailed neurons. J Neuroscience, 44(40).
Proix, T. et al. (2021). Patient-specific network connectivity combined with neural mass models. Front Sys Neurosci.
Marcus, G. (2025). “Scale Is All You Need” is dead. Marcus on AI.
Ghosh, D. & Ghosh, D.P. (2025). The neocortical microcolumn as a memory-retrieval circuit.
Wilkins, A. (2026). We’re about to simulate a human brain on a supercomputer. New Scientist.
EBRAINS. (2026). From brain atlas to personalised model.
Further Reading:
Hopfield, J.J. (1982). Neural networks and physical systems with emergent collective computational abilities. PNAS, 79(8), 2554-2558.
Schirner, M. et al. (2018). Brain network models for multi-scale neurophysiological inference. eLife.
Recent reviews on epilepsy modeling approaches and next generation neural mass models.